












El glucagón es una hormona peptídica producida por las células alfa del páncreas. El glucagón aumenta la concentración de glucosa y ácidos grasos en el torrente sanguíneo y se considera la principal hormona catabólica del cuerpo. El glucagón también se usa como medicamento para tratar una serie de problemas de salud. El efecto del glucagón es opuesto al de la insulina, que reduce la glucosa extracelular. El glucagón se produce a partir del proglucagón, codificado por el gen GCG.
El páncreas libera glucagón cuando la cantidad de glucosa en el torrente sanguíneo es demasiado baja. El glucagón hace que el hígado participe en la glucogenólisis: convierte el glucógeno almacenado en glucosa, que se libera en el torrente sanguíneo. Los niveles altos de glucosa en sangre, por otro lado, estimulan la liberación de insulina. La insulina permite que la glucosa sea absorbida y utilizada por los tejidos insulinodependientes. Por lo tanto, el glucagón y la insulina son parte de un sistema de retroalimentación que mantiene estables los niveles de glucosa en sangre. El glucagón aumenta el gasto energético y se eleva en condiciones de estrés. El glucagón pertenece a la familia de hormonas de la secretina.
Función del glucagón
El glucagón generalmente eleva la concentración de glucosa en la sangre al promover la gluconeogénesis y la glucogenólisis. El glucagón también disminuye la síntesis de ácidos grasos en el tejido adiposo y el hígado. El glucagón promueve la lipólisis en estos tejidos, lo que hace que liberen ácidos grasos a la circulación donde pueden catabolizarse para generar energía en tejidos como el músculo esquelético cuando sea necesario.
La glucosa se almacena en el hígado en forma de glucógeno polisacárido, que es un glucano (un polímero formado por moléculas de glucosa). Las células del hígado (hepatocitos) tienen receptores de glucagón. Cuando el glucagón se une a los receptores de glucagón, las células del hígado convierten el glucógeno en moléculas de glucosa individuales y las liberan en el torrente sanguíneo, en un proceso conocido como glucogenólisis. Cuando el almacenamiento de glucosa se agota, el glucagón estimula al hígado y al riñón a sintetizar glucosa adicional por gluconeogénesis. El glucagón apaga la glucólisis en el hígado, lo que hace que los intermedios glucolíticos sean transportados a la gluconeogénesis.
El glucagón también regula la tasa de producción de glucosa a través de la lipólisis. El glucagón induce la lipólisis en humanos en condiciones de supresión de insulina (como la diabetes mellitus tipo 1).
La producción de glucagón parece depender del sistema nervioso central a través de vías aún por definir. En animales invertebrados, se ha informado que la eliminación del tallo ocular afecta la producción de glucagón. La extirpación del pedúnculo ocular en cangrejos de río jóvenes produce hiperglucemia inducida por glucagón.
Mecanismo de acción del glucagón
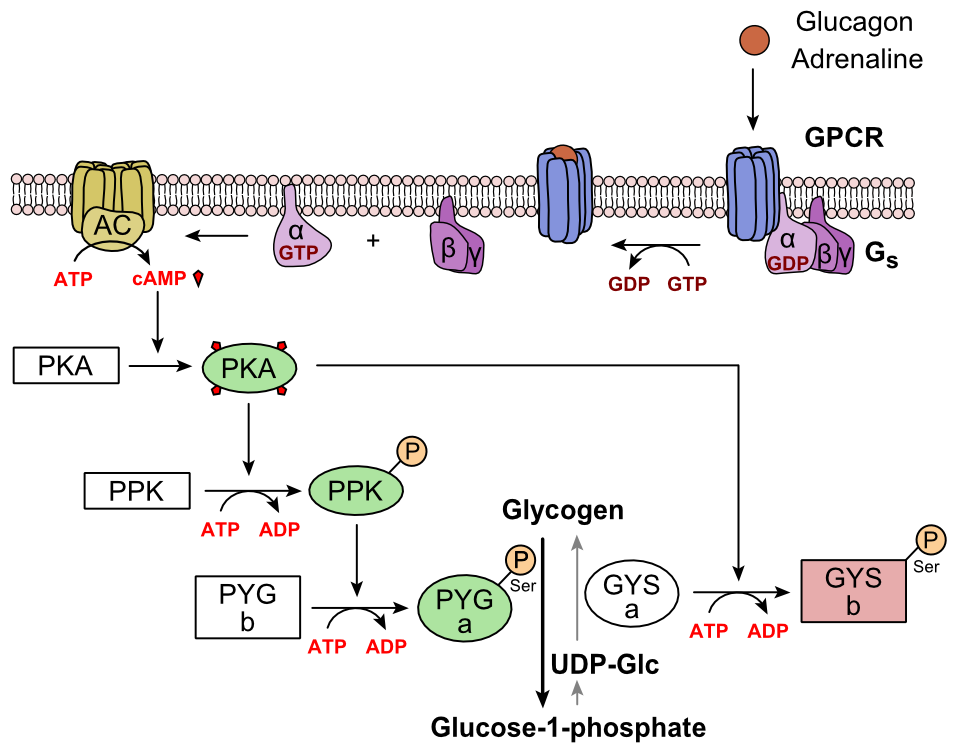
El glucagón se une al receptor de glucagón, un receptor acoplado a proteína G, ubicado en la membrana plasmática de la célula. El cambio de conformación en el receptor activa las proteínas G, una proteína heterotrimérica con subunidades α, β y γ. Cuando la proteína G interactúa con el receptor, sufre un cambio conformacional que resulta en el reemplazo de la molécula de GDP que estaba unida a la subunidad α con una molécula de GTP. Esta sustitución da como resultado la liberación de la subunidad α de las subunidades β y γ. La subunidad alfa activa específicamente la siguiente enzima en la cascada, la adenilato ciclasa.
La adenilato ciclasa fabrica monofosfato de adenosina cíclico (AMP cíclico o AMPc), que activa la proteína quinasa A (proteína quinasa dependiente de AMPc). Esta enzima, a su vez, activa la fosforilasa quinasa, que luego fosforila la glucógeno fosforilasa b (PYG b), convirtiéndola en la forma activa llamada fosforilasa a (PYG a). La fosforilasa a es la enzima responsable de la liberación de glucosa 1-fosfato de los polímeros de glucógeno.
Un ejemplo de la vía sería cuando el glucagón se une a una proteína transmembrana. Las proteínas transmembrana interactúan con Gɑβ𝛾. Gɑ se separa de Gβ𝛾 e interactúa con la proteína transmembrana adenilil ciclasa. La adenilil ciclasa cataliza la conversión de ATP en cAMP. El cAMP se une a la proteína quinasa A y el complejo fosforila la fosforilasa quinasa. La fosforilasa quinasa fosforilada fosforila la fosforilasa. La fosforilasa fosforilada separa las unidades de glucosa del glucógeno como glucosa 1-fosfato.
Además, el control coordinado de la glucólisis y la gluconeogénesis en el hígado se ajusta mediante el estado de fosforilación de las enzimas que catalizan la formación de un potente activador de la glucólisis llamado fructosa 2,6-bisfosfato. La enzima proteína quinasa A (PKA) que fue estimulada por la cascada iniciada por el glucagón también fosforilará un único residuo de serina de la cadena polipeptídica bifuncional que contiene las enzimas fructosa 2,6-bisfosfatasa y fosfofructoquinasa-2. Esta fosforilación covalente iniciada por el glucagón activa al primero e inhibe al segundo. Esto regula la reacción que cataliza la fructosa 2,6-bisfosfato (un potente activador de la fosfofructoquinasa-1, la enzima que es el principal paso regulador de la glucólisis) al disminuir la velocidad de su formación, inhibiendo así el flujo de la vía de glucólisis y permitiendo la gluconeogénesis. para predominar. Este proceso es reversible en ausencia de glucagón (y por tanto, en presencia de insulina).
La estimulación de la PKA con glucagón también inactiva la enzima glucolítica piruvato quinasa en los hepatocitos.
Fisiología
Producción de glucagón
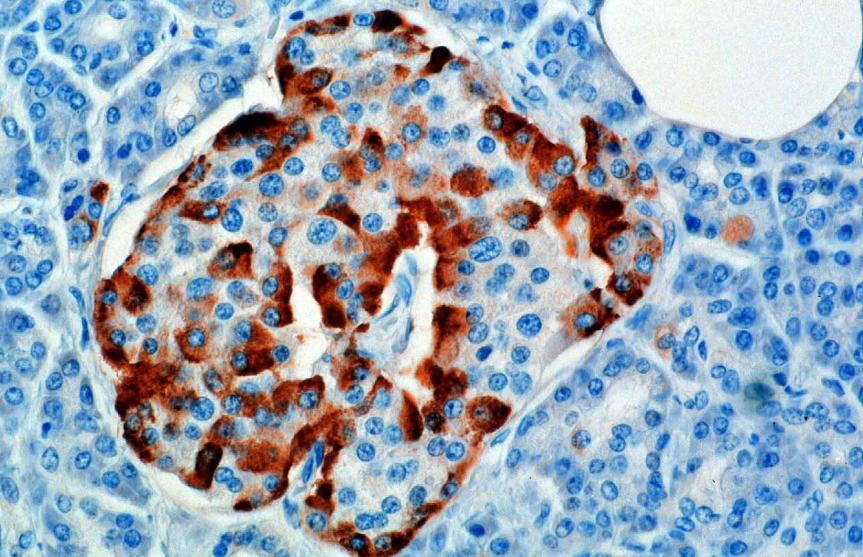
La hormona se sintetiza y secreta a partir de las células alfa (células α) de los islotes de Langerhans, que se encuentran en la porción endocrina del páncreas. La producción, que por lo demás es libre, es suprimida / regulada por la amilina, una hormona peptídica co-secretada con insulina de las células β pancreáticas. A medida que disminuyen los niveles de glucosa plasmática, la reducción subsiguiente de la secreción de amilina alivia la supresión de las células α, lo que permite la secreción de glucagón.
En los roedores, las células alfa se encuentran en el borde exterior del islote. La estructura del islote humano está mucho menos segregada y las células alfa se distribuyen por todo el islote muy cerca de las células beta. El glucagón también es producido por células alfa en el estómago.
Investigaciones recientes han demostrado que la producción de glucagón también puede tener lugar fuera del páncreas, siendo el intestino el sitio más probable de síntesis extrapancreática de glucagón.
Regulación del glucagón
La secreción de glucagón es estimulada por:
- Hipoglucemia
- Epinefrina (a través de los receptores adrenérgicos β2, α2 y α1)
- Arginina
- Alanina (a menudo de la transaminación de piruvato / glutamato derivado del músculo
- Acetilcolina
- Colecistoquinina
- Polipéptido inhibidor gástrico
La secreción de glucagón es inhibida por:
- Somatostatina
- Amilina
- Insulina (a través de GABA)
- Heterodímero del receptor de PPARγ / retinoide X.
- Aumento de los ácidos grasos libres y cetoácidos en la sangre.
- Aumento de la producción de urea
- Péptido-1 similar al glucagón
Estructura del glucagón
El glucagón es un polipéptido de 29 aminoácidos. Su estructura primaria en humanos es: NH2-Su-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp -Leu-Met-Asn-Thr-COOH.
El polipéptido tiene una masa molecular de 3485 daltons. El glucagón es una hormona peptídica (no esteroide).
El glucagón se genera a partir de la escisión del proglucagón por la proproteína convertasa 2 en las células α de los islotes pancreáticos. En las células L intestinales, el proglucagón se escinde en los productos alternativos glicentina, GLP-1 (una incretina), IP-2 y GLP-2 (promueve el crecimiento intestinal).
Patología
Los niveles anormalmente elevados de glucagón pueden ser causados por tumores pancreáticos, como el glucagonoma, cuyos síntomas incluyen eritema migratorio necrolítico, aminoácidos reducidos e hiperglucemia. Puede ocurrir solo o en el contexto de una neoplasia endocrina múltiple tipo 1.
El glucagón elevado es el principal contribuyente a la cetoacidosis hiperglucémica en la diabetes tipo 1 no diagnosticada o mal tratada. A medida que las células beta dejan de funcionar, la insulina y el GABA pancreático ya no están presentes para suprimir la producción de glucagón que corre libremente. Como resultado, el glucagón se libera de las células alfa al máximo, lo que provoca una rápida descomposición del glucógeno en glucosa y una cetogénesis rápida. Se encontró que un subconjunto de adultos con diabetes tipo 1 tardó 4 veces más en promedio en acercarse a la cetoacidosis cuando se les administró somatostatina (inhibe la producción de glucagón) sin insulina. La inhibición del glucagón ha sido una idea popular en el tratamiento de la diabetes, sin embargo, algunos han advertido que hacerlo dará lugar a una diabetes frágil en pacientes con glucosa en sangre suficientemente estable.
Se cree que la ausencia de células alfa (y por lo tanto de glucagón) es una de las principales influencias en la extrema volatilidad de la glucosa en sangre en el contexto de una pancreatectomía total.
Historia
En la década de 1920, Kimball y Murlin estudiaron extractos pancreáticos y encontraron una sustancia adicional con propiedades hiperglucémicas. Describieron el glucagón en 1923. La secuencia de aminoácidos del glucagón se describió a finales de la década de 1950. Una comprensión más completa de su papel en la fisiología y la enfermedad no se estableció hasta la década de 1970, cuando se desarrolló un radioinmunoensayo específico.
.
